Selection Pressure and Artificial Selection in Dogs
Selection pressure is how populations get pushed in one direction rather than another. In dogs, artificial selection means breeders choose which dogs reproduce and which do not. That sounds obvious, but its consequences are often underestimated. Every repeated breeding preference changes allele frequencies over time, and every strong selection target carries a trade-off somewhere else in the genome. Understanding selection honestly is one of the main differences between a breeder who is telling a marketing story and a breeder who is doing population work with awareness of what the population is actually doing in response. Documented
What It Means
The core mechanism
Selection pressure exists whenever some individuals contribute more genes to the next generation than others because of a trait or set of traits. In nature, that process is shaped by survival and reproduction in a given environment. Dogs that survive predators, find food, avoid parasites, and produce viable offspring contribute more to the next generation's gene pool than dogs that do not, and the heritable components of whatever helped them succeed become more common in their descendants. In dog breeding, the same mechanism operates but the criteria are different. Instead of survival and wild reproduction, the filter is human choice. A breeder's preferences decide which dogs reproduce, how often, with whom, and for how many generations. The underlying math is the same. The selection target is different.
Three broad patterns of selection are useful to name because they describe different ways a trait distribution can be pushed over time. Directional selection pushes a trait in one direction. Selecting for larger size, lighter coat color, greater retrieving drive, or lower excitability are all directional moves, because the breeder is consistently favoring dogs at one end of the current distribution and letting the other end drop out of the gene pool over generations. Stabilizing selection favors a middle range and resists extremes on either side. Many breeders do this whether they use the term or not. They want a dog that stays within a preferred window for structure, size, or temperament, and they are quietly pushing back against drift toward either extreme. Disruptive selection favors extremes at both ends and reduces the middle. This pattern is less common in intentional breeding programs, but it can appear when different subpopulations inside the same breed are selecting for sharply different ideal types, which is part of how show and field splits deepen over decades.
Artificial selection is still selection
Artificial selection is simply human-guided selection. The word artificial in this context does not mean fake or unnatural. It means the filter is being applied by human choice rather than by environmental survival pressure. But because it acts inside closed populations, it does more than change one visible trait. It also alters the broader gene-frequency landscape around the selected loci, and it interacts with drift, founder effects, and Ne in ways that make the net outcome harder to predict than a simple "select for X, get more X" mental model would suggest. Documented
The classical breeder's equation captures the main idea conceptually. Response to selection depends on two things: how heritable the trait is in the narrow sense, and how strong the selection differential is (meaning how much the chosen parents differ from the population average on that trait). If a trait has measurable additive genetic variance and breeders consistently favor one end of the distribution, the population mean can move over generations. The expected rate of change per generation is roughly the product of narrow-sense heritability and the selection differential, which is why highly heritable traits respond quickly to selection and why traits with low heritability respond slowly even when breeders try hard.
That is the good news. The breeder's equation tells us that selection does work, that the math is consistent with centuries of empirical breeding progress in dogs and livestock, and that programs applying consistent pressure to measurable traits can genuinely reshape populations on decade timescales.
Why strong selection is rarely free
The caution is that strong selection is rarely free. When breeders select hard for one trait, they often reduce diversity in the genomic regions contributing to that trait because the alleles associated with the favored version become more common while alternatives are squeezed out of the gene pool. Linked loci may come along for the ride because recombination does not perfectly separate neighboring genes within a small number of generations, which means that alleles sitting near the selected locus can rise or fall in frequency alongside it even if nobody was selecting for them directly. This is often called a selective sweep when the pattern is strong, and the population genetics literature on selective sweeps in dog breeds documents them at many loci.
Correlated traits may shift unintentionally as well, because genes often have multiple effects (pleiotropy) or because the selected behavior is tied to a broader neurobiological profile that carries other changes with it. Observed-JB A visible win on the named target trait can carry a hidden cost on traits nobody was tracking, and those costs can take several generations to become obvious because they may be subtle or polygenic themselves.
Dog breeds show this trade-off clearly across many examples. Selection for extreme conformation traits can change far more than outline or coat, often with consequences for airway structure, skeletal loading, or heat tolerance depending on which conformation features were pushed. Observed-JB Selection for working traits can shift arousal baselines, social style, or stress sensitivity alongside task performance, which is why some high-drive working lines produce dogs that are brilliant at their specific job but difficult to live with in a standard pet-home environment. None of that is a moral judgment about show lines or working lines. It is just the reality that genomes are connected systems rather than menus of isolated features, and strong pressure on any feature tends to move neighboring features along with it in ways that are only visible in retrospect.
Selection operates at population scale
This is one reason artificial selection in dogs has to be discussed at the population level rather than the individual level. A single good litter does not mean the population has changed meaningfully. The parents of that litter may have been at the tail end of the current distribution, and their puppies may regress toward the population mean in ways that make them less extreme than the parents were. Regression to the mean is a normal feature of quantitative-trait inheritance and does not indicate that selection has failed; it indicates that the parents were unusual and the offspring partly inherited the usual.
But a decade of consistent selection pressure absolutely can change the population. Across many litters and many generations, even modest selection differentials compound into meaningful shifts in the population mean if the pressure is consistent and the trait is heritable. The breeder's equation is additive across generations in the sense that small per-generation changes accumulate, which is how dog breeds were built in the first place and how they continue to drift or improve depending on which direction their communities are collectively selecting.
Selection inside a small gene pool
Selection inside a closed breed population has one additional complication that selection in a large outbred population does not. The interaction with drift is much stronger. In a small effective population, drift can overwhelm weak selection pressures, so mildly favored alleles may still be lost to random sampling and mildly disfavored alleles may still drift to fixation. The classical breeder's-equation math assumes large-population conditions that closed dog breeds rarely satisfy, which is why the observed response to selection in real breeds often differs from what the clean equation would predict. Observed-JB Selection still works, but the ratio of signal to noise is less favorable than it would be in an infinite or near-infinite population.
JB's position inside this logic
JB's program sits directly inside this logic and is honest about where it sits. The program is not primarily selecting for show-ring flash or maximum working-drive intensity. It is selecting toward a stable, socially legible, low-drama temperament that can live naturally inside the Five Pillars with minimal behavioral management burden for the family. That target is real and it is one the program pursues deliberately across generations. But it is also polygenic, meaning the trait is shaped by many genes of small effect rather than by a single major locus, and polygenic temperament targets respond to selection more slowly and less tidily than highly visible single-trait targets do. Progress on them is visible in trend data across many litters rather than in any one litter's outcome.
That is why careful breeders talk about trend, not magic. Artificial selection can shift the population mean over time. It cannot manufacture a trait overnight or guarantee that every puppy in one litter will perfectly express the target. A breeder who promises that every puppy will have temperament X is either misunderstanding how polygenic inheritance works or is overselling what selection can do on a short timescale. A breeder who says "this is what the line has been trending toward across the last several generations, and most puppies express most of the target most of the time" is describing how selection actually behaves and leaving honest room for the variation that real biology produces.
Why It Matters for Your Dog
What This Cannot Predict
Selection pressure cannot promise perfect outcomes from one mating. The expected shift is a population-level change across generations, not a guarantee for any individual puppy.
It cannot isolate one target trait from the rest of the genome as if there were no correlated costs. Genomes are connected systems, and selection on one feature tends to move neighboring features along with it in ways that can take generations to become visible.
It cannot tell you that strong selection on a trait will always improve the breed overall. Improvement in one domain can narrow the population, pull unwanted correlates along with it, or reduce the diversity available for later selection on different traits.
And it cannot fully escape the interaction with drift in small populations. Even well-targeted selection pressure can fail to produce the expected response when drift is strong enough to overwhelm it.
The correct frame is longer and humbler: selection is powerful across generations, but every strong target requires awareness of the trade-offs it is likely to produce and willingness to measure those trade-offs rather than assume they do not exist.
Families often hear selection language in a vague way. A breeder says they select for health, or structure, or temperament, or working ability. This page helps translate what that really means in population-genetics terms.
It means those traits are not just admired. They are being used to shape the next generation's gene frequencies. Every time the breeder chooses to breed one dog and not another on the basis of a trait, they are applying selection pressure to the allele frequencies associated with that trait, and that pressure will compound across generations whether or not the breeder is consciously tracking the compounding. Documented
That matters because the right breeder questions become more specific and harder to deflect with marketing language. What are you selecting for most strongly, and how strong is that selection actually? How do you know the trait is actually measurable in a way that lets selection work, rather than being too subjective or too environment-dependent for the breeder's equation to apply? What trade-offs are you watching while selecting for it, and how would you notice if an unintended correlated change started showing up? A breeder who can answer those questions is engaged with the real biology. A breeder who cannot is applying selection pressure without understanding what it is doing, which is still selection but it is selection without feedback.
For JB, that last question matters a great deal. Selecting for a calm, thoughtful, socially mature dog is not the same as selecting for a dog that merely looks quiet in one moment. The temperament target has to hold up across line knowledge, adult behavior across many homes, and the way dogs live inside the raising program over time. It has to be measured against the full developmental arc rather than against one observation on one day, because polygenic temperament is noisy and any single snapshot can mislead about the underlying trend. That is a slower but more honest selection project than optimizing for a single observable feature, and the honesty is part of what the Five Pillars framework demands of the program it sits inside.
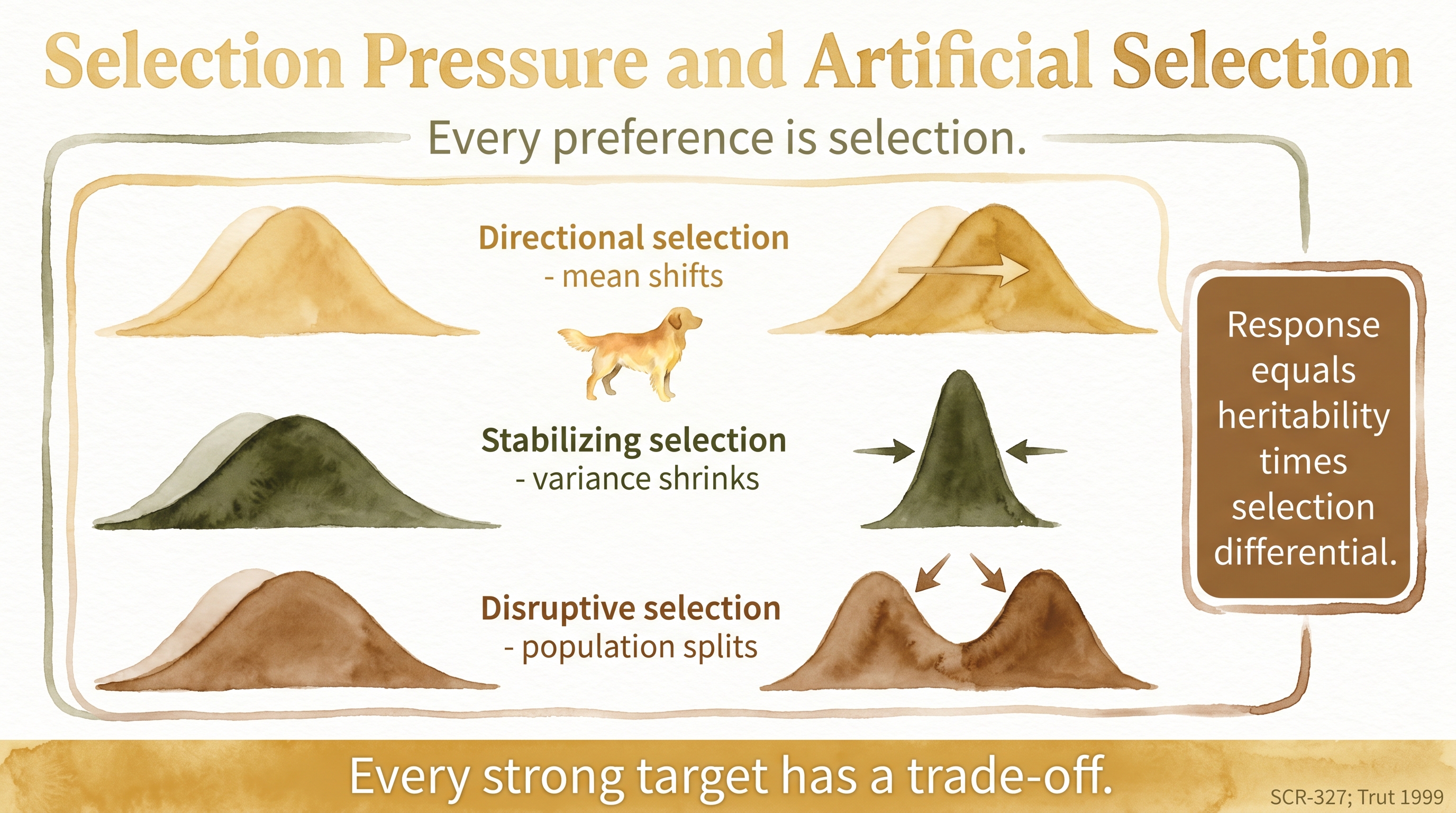
Every repeated breeder preference is selection, and every strong target has a trade-off.
Key Takeaways
- Artificial selection is the human-driven version of selection pressure and works by changing which dogs contribute genes to the next generation.
- Directional, stabilizing, and disruptive selection describe different ways a trait distribution can be pushed over time.
- Strong selection on one trait can reduce diversity and pull correlated traits along with it through linkage and pleiotropy.
- Selection is a long-game population tool: one litter changes little, but years of consistent decisions can reshape a breed segment.
- In closed populations with small Ne, drift can overwhelm weak selection pressures, which is why real response to selection is noisier than the clean equation suggests.
The Evidence
This entry uses observed claim-level tags beyond the dedicated EvidenceBlocks below. These tags mark JB program observation or practice-derived claims that need dedicated EvidenceBlock coverage in a later content pass.
- Quantitative-genetics frameworkgeneral genetics and dogs
Response to selection depends on the heritable component of a trait and the strength of the selection differential applied across generations. - Canine population and behavior literaturedogs
Artificial selection in dogs shifts gene frequencies over time and can alter multiple linked or correlated traits rather than only the one breeders say they are targeting. - Temperament heritability source synthesisdogs
Polygenic behavioral traits can respond to selection, but progress is gradual and sensitive to measurement quality and environmental noise.
- Canine breed-structure literaturedogs
Strong repeated selection on a narrow set of traits can reduce diversity in the contributing genomic regions and may create correlated changes elsewhere in the phenotype. - Canine selective-sweep literaturedogs
Dense genomic marker data across dog breeds documents selective sweeps around loci under strong directional selection, with reduced diversity at linked positions.
No direct measurement exists of what unintended correlated behavioral or physiological changes occurred in Golden Retrievers as a result of selection for specific show or working traits, making it difficult to quantify the full trade-off cost.
SCR References
Sources
- Trut L.N. (1999). Early canid domestication: the farm-fox experiment. American Scientist, 87(2), 160-169.
- Trut L.N., Oskina I., & Kharlamova A. (2009). Animal evolution during domestication: the domesticated fox as a model. BioEssays, 31(3), 349-360.
- Kukekova A.V., Johnson J.L., Xiang X., Feng S., Liu S., Rando H.M., et al. (2018). Red fox genome assembly identifies genomic regions associated with tame and aggressive behaviours. Nature Ecology & Evolution, 2, 1479-1491.
- Wilkins A.S., Wrangham R.W., & Fitch W.T. (2014). The "domestication syndrome" in mammals: a unified explanation based on neural crest cell behavior and genetics. Genetics, 197(3), 795-808.
- Gleeson B., & Wilson L.A.B. (2023). Shared reproductive disruption, not neural crest or tameness, explains the domestication syndrome. Proceedings of the Royal Society B: Biological Sciences. doi:10.1098/rspb.2022.2464
- Bergström A., et al. (2022). Grey wolf genomic history reveals a dual ancestry of dogs. Nature, 607, 313-320.
- Frantz L.A.F., et al. (2016). Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science, 352(6290), 1228-1231.
- Freedman A.H., et al. (2014). Genome sequencing highlights the dynamic early history of dogs. PLOS Genetics, 10(1), e1004016.
- Axelsson E., et al. (2013). The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature, 495, 360-364.