Linkage and Pleiotropy in Canine Genetics
Many dog-genetics misunderstandings begin when people treat an associated marker as if it were the causal gene or treat one visible trait as if it existed in isolation from the rest of the genome. Linkage and pleiotropy explain why that shortcut so often fails in canine genetics. Nearby loci can travel together across generations because recombination does not always separate them, and one gene can influence more than one trait at once because the biology it participates in rarely has only one output. Both mechanisms are fundamental to how genomes actually behave, and both are routinely glossed over in marketing language around genetic tests. Documented
What It Means
Two different mechanisms, one shared consequence
Linkage means loci that sit close together on the same chromosome tend to be inherited together because recombination between them is less likely. Recombination is the process during meiosis in which pairs of chromosomes physically exchange segments, and the further apart two loci are along the chromosome the more often recombination will separate them across generations. Loci that are right next to each other almost always travel together; loci on opposite ends of the same chromosome are separated frequently enough that they behave almost as if they were on different chromosomes. Linkage is a matter of physical distance along the DNA, measured either in base pairs or in centimorgans (the unit used for recombination distance).
Pleiotropy means one gene influences more than one trait. A gene might encode a protein that participates in multiple developmental pathways, or a regulatory region that turns on multiple downstream targets, or an enzyme whose activity affects several phenotypes simultaneously. The classic illustrative example is the gene encoding the melanocortin 1 receptor, which affects coat color in many mammals but also has connections to stress-response systems through its place in a broader signaling network. Documented Pleiotropy is a property of the gene's biology rather than of its chromosomal neighborhood.
Those are different mechanisms, but they create a similar practical problem for anyone trying to interpret a genetic test or make a breeding decision on the basis of a genomic signal: the visible association between a genetic signal and a trait may be more complicated than it first appears. Linkage means the signal you see may be a neighbor of the real cause. Pleiotropy means the real cause may have effects beyond the trait you were looking at. Both limit how confidently any single-marker or single-gene story can be told.
Why linkage matters so much in dog breeds
In dog breeds, linkage is especially important because long linkage-disequilibrium blocks are common. Linkage disequilibrium (LD) is the non-random association of alleles at different loci in a population, and the length of LD blocks in a given breed reflects the breed's history. Breed formation from small founder groups, historical bottlenecks, and strong artificial selection have left dogs with genomic regions in which large chunks of neighboring markers travel together more often than they would in a large, anciently outbred population. Human LD blocks are typically on the order of tens of kilobases. Dog LD blocks within a single breed can extend across several megabases, which is one to two orders of magnitude longer. That is a massive difference, and it has both benefits and costs for canine genetic research.
The benefit is that canine genome-wide association studies (GWAS) can work with smaller sample sizes than many human studies. Documented-Cross-Species If large linked blocks are moving together, a marker can tag a disease-associated region without sitting directly on the causal mutation, and a modest number of dogs can be enough to pinpoint which block of the genome is associated with the trait. Several canine disease genes were mapped with sample sizes that would have been hopelessly underpowered in a human GWAS, because the long LD blocks in breeds made the statistical signal much easier to detect. Canine genetics has exploited this advantage aggressively over the last two decades and it is one of the reasons the field has progressed so quickly.
The cost of long LD blocks
But that same strength creates a caution that becomes critical when the results get translated into breeder-facing tests. A marker associated with a disease is not necessarily the causal variant. It may simply be traveling near the causal variant in the breed where the study was performed, sitting somewhere inside the same long LD block. The marker tags the block; the causal mutation lives somewhere inside the block but might be tens or hundreds of thousands of bases away from the marker itself. Within the original breed, the marker works well as a diagnostic because the marker allele and the causal allele almost always travel together in that population.
The problem emerges when the marker is used in a different breed. Different breeds have different LD structures because their histories and recombination events differ. A marker that is tightly linked to a causal variant in Breed A may not be linked to it in Breed B at all, because the historical recombination events in Breed B have separated them. The marker may still appear on the dog's chip and may still return a result, but the result has no predictive value in the new breed because the association the test was built on does not hold.
That is why some marker-based tests work well in one breed and fail in another. The marker was validated in the original population's linkage structure. Change the breed background, recombination history, or local haplotypes, and the association may weaken or disappear. This is a well-documented problem in canine genetic testing, and it is one reason the literature repeatedly emphasizes that breed-specific validation is required before a test developed in one breed should be marketed to another. Commercial panels that ignore this principle and sell the same marker across many breeds are overclaiming what the underlying science supports.
Pleiotropy and correlated consequences
Pleiotropy creates a different but equally important issue. Sometimes a single gene truly does affect multiple traits, not because of linkage to other genes but because the gene's own biological function has multiple downstream consequences. That means selection on one trait can unintentionally move another because the same biological pathway is involved. In dogs, this is why highly visible traits such as coat pattern or morphology can come with broader consequences than the casual breeder expects.
Classic pleiotropic examples in dogs include loci affecting coat color that also appear to influence behavioral or physiological traits in the same animals. The color locus is not "controlling" the behavior in any deep causal sense; it is connected to the same signaling network, and changes at the color locus can ripple through the network to produce downstream effects on other phenotypes. Selection focused on the visible color may be dragging the less-visible correlates along with it whether or not the breeder intended that outcome.
Pleiotropy can also work in the other direction. A gene with real medical significance may have a cosmetic side effect that breeders notice, and selection on the cosmetic side effect may accidentally be selecting on the medical trait without anyone realizing it. This is one of the more insidious failure modes of naive selection programs because the breeder is not even aware that a medically important trait is being shaped by their decisions.
The connected-genome framing
The practical lesson from both mechanisms is that genomes are not organized around human convenience. Visible traits, disease risk, behavior, and physiology are not isolated boxes that can be selected on independently. Linkage means regions travel together whether you want them to or not. Pleiotropy means one gene can do more than one job whether you know about the other jobs or not. A breeder who treats each trait as a separable variable in a selection program is using a mental model that does not match how the underlying biology works.
That makes single-gene storytelling especially risky in dogs. A breeder may say a marker is "for" a trait, when what the study actually found was a statistical association in a linked region that may or may not hold up in the specific dog being tested. Documented Or a breeder may select hard on one aesthetic feature without realizing that a pleiotropic or tightly linked effect is being carried along at rates that will become visible only several generations later. Both mistakes look fine in the moment and become expensive in the long run, which is why understanding linkage and pleiotropy is not an academic exercise but a practical requirement for honest program management.
Why It Matters for Your Dog
What This Cannot Predict
An associated marker does not mean the dog will get the disease. Association is a statistical statement about populations, not a deterministic statement about individuals, and even a strong association leaves room for many individual dogs to not match the population trend.
A nearby signal in a GWAS does not automatically identify the causal gene. The GWAS signal points to a genomic neighborhood; identifying the specific causal variant usually requires additional fine-mapping work, functional studies, and replication in independent samples. Observed-JB
A pleiotropic effect found in one context does not always tell you the exact magnitude of the effect in every breed. Documented Pleiotropic relationships can vary across genetic backgrounds because the networks they participate in differ in their other components.
And a marker validated in one breed may not work in another, or may work with a different strength, or may return results that look meaningful but are uninformative because the underlying LD structure has changed.
This is why families should be cautious whenever test results are marketed with more certainty than the underlying association warrants. "Associated with" and "causes" are not interchangeable phrases, and collapsing them is one of the most common ways canine genetic testing gets oversold to the public.
This page matters because it helps families and breeders ask a more intelligent question about genetic tests: what exactly is this test measuring at the molecular level, and how confident is the chain of reasoning from the molecular signal to the clinical claim?
If the answer is a direct causal mutation that has been functionally validated and mechanistically understood, interpretation may be strong and the test result can be treated with confidence. Several canine disease tests do meet this standard, and they are among the most useful tools in modern breeding.
If the answer is a marker linked to a disease region but not the causal variant itself, the interpretation is more conditional and breed-dependent. The test may be informative in the breed where it was validated and essentially random noise in a breed it has not been validated in, and the marketing around the test may or may not make that distinction clearly.
If the answer is a polygenic score built from many associated regions across the genome, the interpretation is even more probabilistic and should be framed as a risk estimate rather than a diagnostic result. Polygenic scores in dogs are an active research area and are valuable directionally but should not be read as individual-level predictions with the confidence that single-gene tests deserve.
Linkage and pleiotropy are the reasons those categories need to stay separate in conversation rather than being collapsed under the single label of "genetic test." For breeders, the lesson is practical: selection on one trait may carry correlated consequences through linked regions or shared biological pathways, and those consequences will emerge in the program whether or not the breeder was watching for them. For families, the lesson is interpretive: a marker result is not a guarantee simply because it sounds genomic, and the appropriate level of confidence depends on which category of test produced it and whether the test was validated in the specific breed being tested.
For JB specifically, this matters because honest communication with families requires distinguishing between the tests the program relies on because they are directly causal and validated, and the tests the program treats more cautiously because they are association-based or breed-transfer-dependent. Collapsing those distinctions in conversation would oversell the certainty of the program's screening, which is exactly the kind of slippage the Five Pillars framework and the JB evidence standards are designed to prevent.
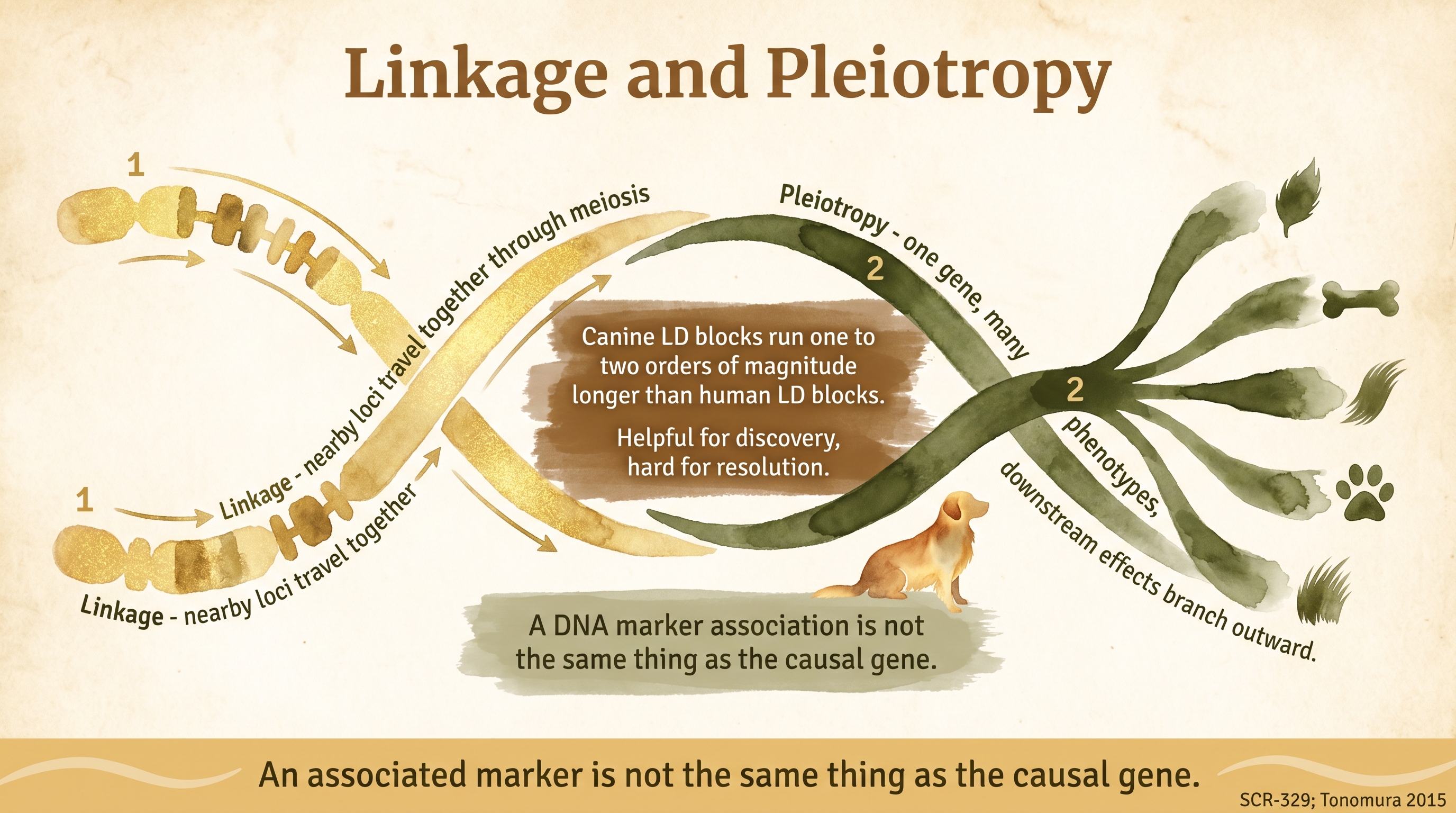
An associated marker is not the same thing as the causal gene.
Key Takeaways
- Linkage means nearby loci can travel together, so an associated marker is not always the causal variant.
- Dog breeds have long linkage-disequilibrium blocks, which makes canine GWAS useful but also broad rather than surgically precise.
- Pleiotropy means one gene can affect multiple traits, so selection on one visible feature can shift others unintentionally.
- In canine genetics, associated with and causes are not interchangeable claims.
- A marker validated in one breed may fail in another because LD structure differs across breed histories, which is why breed-specific validation matters.
The Evidence
This entry uses documented cross-species claim-level tags beyond the dedicated EvidenceBlocks below. Species and application scope should be checked during the next evidence-chain authoring pass.
This entry uses observed claim-level tags beyond the dedicated EvidenceBlocks below. These tags mark JB program observation or practice-derived claims that need dedicated EvidenceBlock coverage in a later content pass.
- Population-genetics frameworkgeneral genetics
Linked loci on the same chromosome are inherited together more often when recombination between them is infrequent. - Canine genomics literaturedogs
Dog breeds often show long linkage-disequilibrium blocks because of bottlenecks, founder effects, and artificial selection, making association mapping both powerful and potentially coarse. - Canine disease-testing literaturedogs
Marker-based tests can depend heavily on breed-specific linkage structure and are not automatically transferable across populations.
- Canine and comparative genetics literaturedogs and other species
Some genes influence more than one trait, meaning selection on a visible or desired trait can bring unintended biological correlates along with it. - Canine coat-color and behavior literaturedogs
Some coat-color and morphology loci in dogs show pleiotropic relationships with other phenotypes, illustrating that selection on visible traits can carry correlated effects.
No comprehensive study has mapped which specific coat-color or morphology loci in Golden Retrievers carry pleiotropic behavioral effects, or measured the magnitude of those correlations in the breeding population.
SCR References
Sources
- Tonomura N., Elvers I., Thomas R., Megquier K., Turner-Maier J., Howald C., et al. (2015). Genome-wide Association Study Identifies Shared Risk Loci Common to Two Malignancies in Golden Retrievers. PLOS Genetics, 11(2), e1004922. doi:10.1371/journal.pgen.1004922
- Ivansson E.L., Megquier K., Kozyrev S.V., Murén E., Körberg I.B., Swofford R., et al. (2016). Variants within the SP110 nuclear body protein modify risk of canine degenerative myelopathy. Proceedings of the National Academy of Sciences, 113(22), E3091-E3100. doi:10.1073/pnas.1600084113
- Bannasch D., Famula T., Donner J., Anderson H., Honkanen L., Batcher K., Safra N., Thomasy S., & Rebhun R. (2021). The effect of inbreeding, body size and morphology on health in dog breeds. Canine Medicine and Genetics, 8(1), 12. doi:10.1186/s40575-021-00111-4
- Wilkins A.S., Wrangham R.W., & Fitch W.T. (2014). The "domestication syndrome" in mammals: a unified explanation based on neural crest cell behavior and genetics. Genetics, 197(3), 795-808.
- Hill W.G. & Weir B.S. (2011). Variation in actual relationship as a consequence of Mendelian sampling and linkage. Genetics Research, 93(1), 47-64.
- Pfahler S., Distl O., & ARCA Members. (2015). Effective Population Size, Extended Linkage Disequilibrium and Runs of Homozygosity in the Norwegian Lundehund. PLoS ONE, 10(4), e0122680. doi:10.1371/journal.pone.0122680