Assortative Mating and Non-Random Mating in Dog Breeds
No dog breed mates randomly. Breeders choose pairings deliberately, and they often choose dogs that resemble each other in the traits they value most. That pattern is called assortative mating. It sounds technical, but it describes one of the most ordinary facts of breeding practice: type seeks type, and that has population-genetic consequences that compound across generations whether or not anyone is tracking them. The random-mating assumption that underpins most introductory population-genetics math is never satisfied by any functioning dog breed, and the distance between the idealization and the reality is where a lot of the hidden diversity loss in closed populations actually happens. Documented
What It Means
The three mating structures
Assortative mating means pairing individuals that are more similar to each other than random chance would produce.
Positive assortative mating means like is being bred to like. A breeder chooses dogs similar in structure, style, pedigree background, or temperament, because they are trying to reproduce a recognizable type and they believe the best way to do that is to use parents who already embody the type. Documented This is by far the most common pattern in purebred dog breeding, and it is the one most programs use as their default whether or not they call it assortative mating.
Disassortative mating means deliberately pairing dogs that differ in a way intended to widen or balance the next generation. A breeder might pair a dog with a refined head to one with a heavier build to shoot for a middle outcome, or pair a very driven dog to a calmer one to soften arousal in the offspring. Documented Disassortative mating is less common as a default but appears in programs that are specifically trying to restore balance or expand a trait range.
Random mating is the Hardy-Weinberg idealization in which every dog has an equal probability of mating with every other dog, independent of any trait or pedigree relationship. It is useful as a null model in population-genetics calculations because it produces clean math, and many derivations start from the random-mating assumption to keep the equations tractable. But it is not how dog breeding works in reality, and most of the population-genetics intuitions that assume random mating need to be adjusted when they are applied to actual breed populations.
Why positive assortative mating is so common
Positive assortative mating is common because it helps breeders reproduce a recognizable type. If a breeder values a specific outline, coat, movement pattern, or social style, they often choose dogs that already share those characteristics rather than gambling on a cross that might produce puppies who look and behave very differently from either parent. Observed-JB The instinct is reasonable. Predictability in breeding output is a legitimate goal, both for the breeder's planning and for the families who are hoping to receive the kind of dog the breeder described to them.
That can make phenotypes more predictable in the short run, which is one of the main benefits breeders are buying with the strategy. It can also concentrate the underlying genetics more quickly than casual pedigree review suggests, which is the less-discussed cost. Documented When two dogs resemble each other closely in multiple traits, they often resemble each other genetically in the regions that produce those traits, even if their immediate pedigree relationship does not look unusually close on a four-generation pedigree chart. Pedigree COI calculated over a short window will not capture the similarity because the relatedness may live in older shared ancestors that fall outside the pedigree window, and genomic COI based on dense marker data will usually show higher values than the pedigree estimate in those cases.
Hidden relatedness and the same regions twice
This is important because repeated similarity is not only about visible type. It often overlaps with hidden relatedness in the very genomic regions the breeder is selecting on. Two dogs may come from similar lines, carry similar haplotypes in the regions contributing to the traits under selection, and reinforce the same concentrated genomic patterns even if their immediate pedigree relationship does not look unusually close. The breeder is effectively compounding line concentration at the loci they care about most, which is the opposite of the diversity preservation the pedigree chart would suggest.
That is why assortative mating can raise homozygosity at selected loci faster than a simple short-generation COI might imply. The breeder is not only choosing relatedness in an abstract sense. They are choosing similarity in the very regions they care most about preserving, and those regions accumulate homozygosity faster than the genome-wide average because every mate choice is applying selection pressure in the same direction at those specific positions. Genome-wide COI averages across the whole genome and smooths out local concentration; assortative mating produces concentration that is local rather than global, which genome-wide averages can miss.
The consequence is that a breeder can look at their pedigree chart and conclude the line is well managed because the displayed COI is modest, while the genomic regions under selection are actually becoming quite concentrated. The discrepancy between pedigree-level and genome-level metrics is exactly what genomic COI was developed to detect, and it often reveals that programs relying on visible type matching have more local homozygosity than their paper pedigrees would suggest.
The popular sire as an extreme case
The popular-sire effect is an extreme form of non-random mating. When one male contributes an outsized number of offspring, breeders are not just choosing like with like across many independent pairings. They are also saturating the population with one paternal contribution, which affects every line that uses the sire and every downstream generation that descends from those lines. That changes allele frequencies, narrows effective population size, and reduces future flexibility in ways that compound much faster than distributed mate choice would.
Popular sires arise for several interlocking reasons. A male who produces a well-regarded litter becomes more attractive as a stud. Breeders who want to reproduce that kind of output book him repeatedly, which produces more well-regarded litters, which attracts more bookings. The dynamic is self-reinforcing across several breeding seasons, and by the time the breed community recognizes that one male's contribution has become disproportionate, the damage to gene-pool balance is already distributed across many programs and many puppies. Popular-sire episodes are one of the main mechanisms by which breeds lose effective population size even when their census counts look healthy.
Golden Retrievers illustrate this problem well because the breed is large enough to look genetically comfortable on census metrics while still allowing relatively narrow sire concentration to shape a great deal of the next generation. That is one reason population-level metrics can look more concerning than breed popularity would suggest when analyzed honestly. The breed is not small in headcount. It is structurally concentrated in ways that the headcount cannot see.
Disassortative mating as a corrective tool
Disassortative mating can sometimes help restore breadth in a program that has grown too concentrated. Breeding across more distant lines inside the breed can interrupt excessive concentration and widen the next generation's options at loci where homozygosity has been accumulating. Programs using this strategy deliberately might pair a heavily linebred bitch with a male from a less-related line, accepting some variability in the resulting puppies as the price of restoring diversity to the gene pool.
But even that strategy works only when the breeder understands what is actually being widened. Pairing visible differences while keeping the same hidden line concentration does less than people think. If both dogs still trace back to the same handful of influential ancestors a few generations earlier, the disassortative appearance of the pairing is cosmetic and the actual gene pool continues to narrow. Honest disassortative mating requires looking past phenotype differences to the underlying pedigree and, where possible, to the underlying genomics.
Disassortative mating also has its own costs that breeders should be honest about. It produces more variable litters, which can be harder to market and harder to place into homes expecting specific outcomes. It can temporarily interrupt the type consistency the program has been building. It takes longer to pay off in visible results because the diversity gains are measured in structural metrics rather than in show-ring successes. Documented None of those costs make it a bad strategy. They just mean the decision to use it should come with clear eyes about what it is trading.
Why It Matters for Your Dog
What This Cannot Predict
Assortative mating is not automatically bad. It is the normal strategy most breeders use, and in moderation it is consistent with responsible population management when it is balanced against diversity awareness.
It is not a synonym for irresponsible breeding. Breeders using positive assortative mating carefully and tracking their line concentration are not doing the same thing as breeders who are recklessly concentrating a program around a single fashionable line.
And it does not mean every type-consistent breeder is damaging the breed. Some level of type consistency is the whole point of having breeds in the first place, and consistency requires some degree of assortative mating.
The point is that positive assortative mating has a cost structure as well as a benefit structure, and both need to be visible when the strategy is being used. It can stabilize desired traits, which is valuable. It can also concentrate the same genomic regions again and again, which is costly. The right question is not whether it exists in a program. The right question is whether the breeder understands its long-term population consequences and balances it against diversity-aware counterpressures.
Families often admire consistency in a breeder's dogs, and consistency can indeed reflect thoughtful selection and careful mate pairing over many years. But consistency is never free. It comes from repeated population choices that are also gradually narrowing the pool those dogs come from.
That means a buyer can ask more useful questions than "do your puppies look the same?" How wide is the breeding pool behind this consistent type, and are you tracking its breadth across generations? Are the same male lines being used repeatedly across your litters, or is there meaningful variety in who is contributing? How do you preserve diversity while still selecting for your preferred temperament and structure, and what specific moves in your program reflect that diversity awareness? A breeder who can answer those questions has thought about assortative mating as a real variable in their population work. A breeder who treats every pairing as independent from every other pairing is applying assortative mating without tracking its cumulative effect.
For JB, this matters because a stable temperament target is valuable, and positive assortative mating is part of how any temperament-focused program actually produces consistent outcomes. But the program cannot pursue that target by acting as though the same narrow line concentration is harmless indefinitely. Good breeding balances type preservation with enough breadth to keep the population biologically flexible for the next decade of work, not just the current litter. The Five Pillars framework depends on puppies having the physical and biological substrate to express the temperament they were selected toward, and that substrate is exactly what diversity management protects. Assortative mating used without diversity awareness eventually eats into that substrate and costs the program the very trait it was trying to preserve.
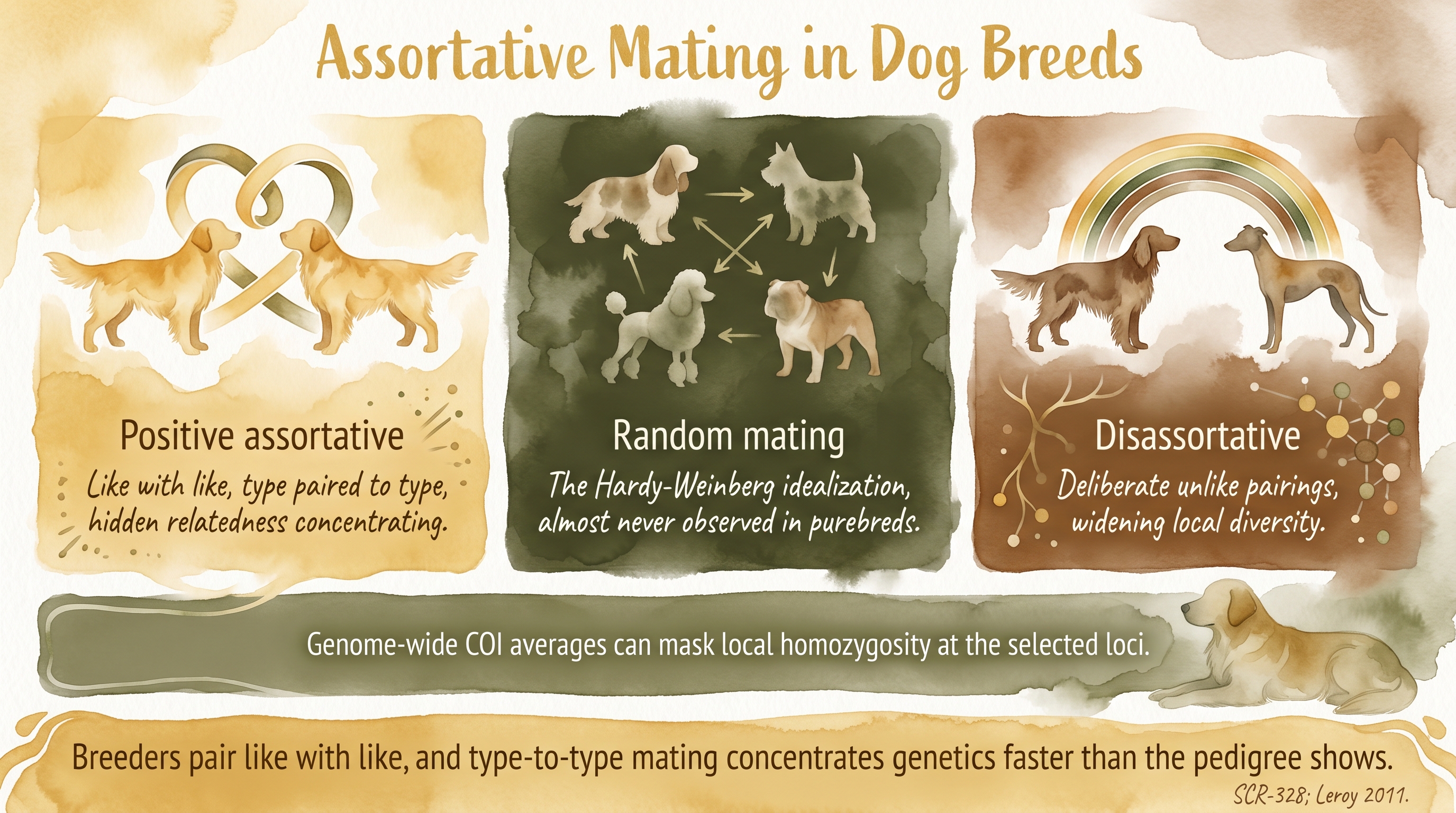
Breeders pair like with like, and type-to-type mating concentrates genetics faster than the pedigree shows.
Key Takeaways
- Dog breeds do not mate randomly; most breeding programs use some degree of assortative mating whether they name it or not.
- Positive assortative mating helps preserve a type, but it can also concentrate homozygosity in the regions under selection.
- The popular-sire effect is an intensified form of non-random mating with major consequences for diversity.
- Good breeders do not avoid assortative mating entirely; they manage its long-term costs instead of pretending they do not exist.
- Short-window pedigree COI can miss the local homozygosity produced by assortative mating at selected loci, which is where genomic COI becomes useful.
The Evidence
This entry uses observed claim-level tags beyond the dedicated EvidenceBlocks below. These tags mark JB program observation or practice-derived claims that need dedicated EvidenceBlock coverage in a later content pass.
- Population-genetics frameworkgeneral population genetics
Random mating is a theoretical baseline, while assortative and disassortative mating describe the real pairing structures seen in managed populations. - Canine breeding-structure literaturedogs
Positive assortative mating is common in purebred dogs because breeders repeatedly pair animals similar in the traits they are trying to preserve. - Canine demographic studiesdogs
When repeated similarity overlaps with narrow sire use and line concentration, homozygosity and effective population narrowing can rise faster than casual pedigree review suggests.
- Canine diversity literaturedogs and Golden Retrievers
The popular-sire effect is an extreme form of non-random mating that distorts allele frequencies and contributes to long-term diversity loss. - Pedigree versus genomic COI comparisonsdogs
Pedigree COI calculated over short windows often understates the local homozygosity produced by repeated assortative mating at selected loci, which genomic COI based on dense marker data can reveal.
No published study has directly quantified the long-term diversity loss from assortative mating in Golden Retrievers specifically across controlled breeding timescales, which would definitively measure the population-genetic cost of type-matching strategies.
SCR References
Sources
- Leroy G. (2011). Genetic diversity, inbreeding and breeding practices in dogs: Results from pedigree analyses. The Veterinary Journal, 189(2), 177-182. doi:10.1016/j.tvjl.2011.06.016
- Leroy G., Phocas F., Hedan B., Verrier E., Elsen J.M., & Mangin B. (2015). Inbreeding impact on litter size and survival in selected canine breeds. The Veterinary Journal, 203(1), 74-78. doi:10.1016/j.tvjl.2014.11.008
- Windig J.J. & Oldenbroek K. (2015). Genetic management of Dutch golden retriever dogs with a simulation tool. Journal of Animal Breeding and Genetics, 132(6), 428-440. doi:10.1111/jbg.12149
- Calboli F.C.F., Sampson J., Fretwell N., & Balding D.J. (2008). Population structure and inbreeding from pedigree analysis of purebred dogs. Genetics, 179(1), 593-601. doi:10.1534/genetics.107.084954
- Wijnrocx K., Bekaert M., Georges M., & Leroy G. (2016). Half of 23 Belgian dog breeds has a compromised genetic diversity, as revealed by genealogical and molecular data analysis. Journal of Animal Breeding and Genetics, 133(5), 375-383. doi:10.1111/jbg.12203